被子植物的生命周期中,從營養(yǎng)生長到生殖生長(開花)的轉(zhuǎn)變是一個至關(guān)重要的發(fā)育過程,也是農(nóng)作物的重要農(nóng)藝性狀,成花轉(zhuǎn)變直接影響著農(nóng)作物的產(chǎn)量和質(zhì)量。冬小麥為主要糧食作物,廣泛種植于溫帶地區(qū)。冬麥在秋天播種,以幼苗越冬,需要經(jīng)歷長時間的持續(xù)低溫(冬季低溫)才能獲得在來年春季開花的潛力(春化作用)。春化作用賦予冬小麥“越冬記憶”,確保春季氣溫變暖后,植物在營養(yǎng)生長后期及時轉(zhuǎn)變至生殖生長(開花),但開花結(jié)實后產(chǎn)生的下一代又需要越冬才能獲得開花的潛力。這樣,每一代冬小麥都需要經(jīng)歷冬季持續(xù)低溫,才能在春天開花結(jié)實。
近日,何躍輝團隊在NATURE PLANTS發(fā)表了題為“A molecular mechanism for embryonic resetting of winter memory and restoration of winter annual growth habit in wheat”的研究文章,揭示了冬小麥“越冬記憶”重置與越冬習性在子代復原的分子機制。

春化通路在不同種類植物(不同的科)的進化過程中獨立形成,因此,不同春化通路的核心基因及分子與表觀遺傳調(diào)控機理迥異。例如,在十字花科的擬南芥中,春化通路響應(yīng)持續(xù)低溫,沉默核心開花抑制基因FLC的表達,從而使植物能在春季開花結(jié)實,但在溫帶谷類作物中(如小麥、大麥、黑麥),VRN1-VRN2-VRN3 (VERNALIZATION 3) 組成了春化的核心調(diào)控網(wǎng)絡(luò)1,2。VRN1及VRN2的激活與抑制決定了小麥的春、冬性。在春化前(未經(jīng)低溫處理)的麥苗中,促進成花轉(zhuǎn)變的核心轉(zhuǎn)錄因子VRN1的表達處于沉默狀態(tài),而抑制成花轉(zhuǎn)變的轉(zhuǎn)錄因子VRN2的表達處于激活狀態(tài),防止小麥在越冬時開花;持續(xù)低溫處理逐漸激活VRN1表達,并抑制VRN2的表達,進而在春化(冬季)結(jié)束后,高水平的VRN1蛋白激活VRN3表達(VRN3為擬南芥FT的同源基因,編碼促進開花的成花素蛋白),最終促進小麥進入生殖生長(開花)。冷誘導的VRN1表達激活狀態(tài)及VRN2抑制狀態(tài)在春天升溫后的植物生長發(fā)育過程中得以維持,形成“越冬記憶”,但這種記憶或春化狀態(tài)也必須在某一時期以某種方式進行重置以保證子代植株同樣需要經(jīng)歷寒冬(春化)后才能開花。
何躍輝課題組的最新研究發(fā)現(xiàn),持續(xù)低溫誘導的VRN1激活狀態(tài)可以遺傳至早期胚胎,在隨后的胚胎發(fā)育過程中逐漸被重置至沉默狀態(tài),而VRN2的表達在胚胎發(fā)育過程中一直處于沉默狀態(tài),但在子代小麥種子萌發(fā)過程中,被多種波長的光快速激活,因此,在幼苗中VRN1處于沉默狀態(tài),而VRN2處于激活狀態(tài),從而在子代重新建立了越冬習性,即又需要經(jīng)歷寒冬誘導的VRN1表達激活及VRN2的沉默,小麥才能在春天開花結(jié)實。此外,該研究發(fā)現(xiàn)在小麥組培再生過程中,VRN2的表達在葉片中被光信號訊速激活,因此,源自冷處理親本的再生小麥植株也需經(jīng)歷春化,才能獲得開花潛力。該研究揭示了冬小麥“越冬記憶”重置與冬性重建的兩步分子機制:VRN1在胚胎發(fā)育過程中被沉默,VRN2在種子萌發(fā)后被光信號激活。
該研究進一步發(fā)現(xiàn)VRN1位點的表觀標記-組蛋白甲基化修飾(H3K4me3、H3K36me3和H3K27me3)在春化過程及小麥世代交替中與其抑制/激活狀態(tài)密切相關(guān)。在此基礎(chǔ)上,研究者對小麥中一個H3K27me3閱讀蛋白RVR1進行突變,從而證實了染色質(zhì)修飾在調(diào)控小麥發(fā)育及春化過程中對VRN1的重要調(diào)控作用。基于這些發(fā)現(xiàn),作者提出了一個持續(xù)低溫及發(fā)育信號調(diào)控VRN1表達的工作模型(圖1)。在寒冬來臨之前,H3K27甲基化酶復合體PRC2催化VRN1染色質(zhì)的H3K27me3修飾,閱讀蛋白RVR1與其它轉(zhuǎn)錄抑制性因子協(xié)同抑制VRN1表達,這與VRN2的表達激活,共同賦予了小麥的越冬生長習性。冬季的長時間寒冷(持續(xù)低溫)導致VRN1染色質(zhì)的H3K27me3大幅減少,進而富集了轉(zhuǎn)錄激活性的H3K4me3和H3K36me3;此外,低溫還通過冷響應(yīng)的轉(zhuǎn)錄調(diào)控因子誘導VRN1的表達。因此,在春化過程中,長時間持續(xù)低溫處理幼苗會導致VRN1染色質(zhì)狀態(tài)從抑制轉(zhuǎn)變?yōu)榧せ睢H欢陂_花受精后的胚胎發(fā)育過程中,VRN1染色質(zhì)狀態(tài)又會切換回抑制狀態(tài)。
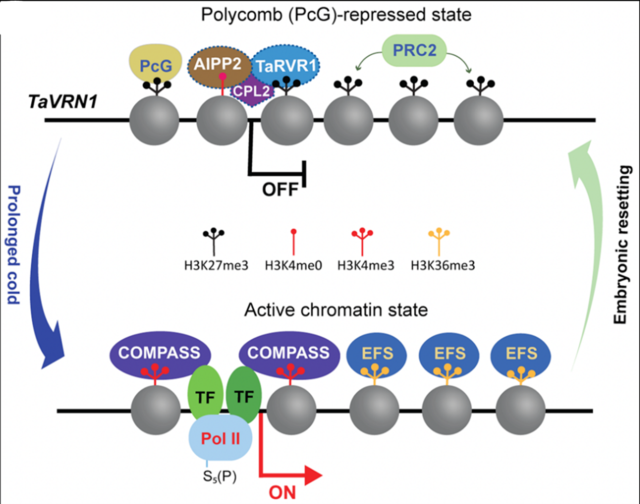
圖1 表觀修飾因子與轉(zhuǎn)錄因子協(xié)同介導長期低溫和發(fā)育信號調(diào)控VRN1在小麥生命周期不同階段的表達 (https://doi.org/10.1038/s41477-023-01596-6)3。
總之,這項研究闡明了冬小麥“越冬記憶”重置與越冬習性在子代復原的兩步分子機制,加深了我們對冬小麥春化過程的理解。同時,該研究也為遺傳改良小麥的冬性提供了新的基因靶點。
北京大學現(xiàn)代農(nóng)業(yè)研究院的牛德副研究員和北京大學現(xiàn)代農(nóng)學院的高政博士為共同第一作者,北京大學現(xiàn)代農(nóng)學院的研究生崔博文及現(xiàn)代農(nóng)業(yè)研究院的張永興博士為共同作者,北京大學現(xiàn)代農(nóng)學院的何躍輝教授為通訊作者。該研究得到了山東省重點研發(fā)計劃、國家自然科學基金及北京大學-清華大學生命科學聯(lián)合中心相關(guān)經(jīng)費的資助。
參考文獻
1. Chen, A., and Dubcovsky, J. (2012). Wheat TILLING mutants show that the vernalization gene VRN1 down-regulates the flowering repressor VRN2 in leaves but is not essential for flowering. PLoS Genet 8, e1003134.
2. Xu, S., and Chong, K. (2018). Remembering winter through vernalisation. Nat. Plants 4, 997-1009.
3. Niu, D., Gao, Z., Cui, B., Zhang, Y. and He, Y. A molecular mechanism for embryonic resetting of winter memory and restoration of winter annual growth habit in wheat. (2024) Nat. Plants, https://doi.org/10.1038/s41477-023-01596-6.